
Les associations entre espŤces du rťcif
Date: 03 aoŻt 2005 à 00:00:00 CEST
Sujet: Thťorie

La relation la plus connue qui lie les espŤces d'un mÍme ťcosystŤme est la prťdation. Cependant, il existe de nombreux autres types de relations interspťcifiques: le mutualisme, le parasitisme, le commensalisme Ö Ces diffťrentes relations sont classťes en fonction des comportements trophiques - c'est ŗ dire la maniŤre de se nourrir - par rapport aux coŻts et bťnťfices quíen retirent les deux partenaires. Tous les modes d'associations se retrouvent sur le rťcif, et on observe un certain nombre en aquarium.
Les diffťrentes modes d'interactions
La synoecie
Cíest lorsquíil níy a ni bťnťfice ni prťjudice entre les deux espŤces, la relation est neutre. L'espace et les ressources sont partagťs. On parle aussi de cohabitation.
La prťdation
La prťdation se dťfinit par líinteraction entre une espŤce dite prťdatrice qui consomme une autre espŤce dite proie. Dans le milieu naturel, cette relation síťquilibre d'elle-mÍme. Le nombre des prťdateurs dťpend directement du nombre de proies, la population des prťdateurs restant faible en gťnťral. Dans líťcosystŤme quasi fermť de nos aquariums, il existe rarement un tel ťquilibre. Cíest pour cela que líintroduction de prťdateurs spťcifiques de parasites - cíest ŗ dire dont le rťgime alimentaire est spťcialisť, comme les crevettes arlequins Hymenocera sp. pour lutter contre les asterines Asterina sp. par exemple - fonctionne rarement, líespŤce prťdatrice disparaissant avant líespŤce prťdatťe. Dans le cas des Hymenocera, il faudrait Ítre en mesure de fournir continuellement des ťchinodermes en quantitť suffisante pour les nourrir (un clichť d'Hymenocera picta).


Asterina sp.
Photos, Christian SEITZ
On observe la mÍme chose avec les nudibranches Berghia verrucicornis pour lutter contre les anťmones de verre Aiptasia ou Chelidonura varians qui consomme les planaires parasites Convolutriloba retrogemma (quand ces limaces de mer ne sont font pas tout simplement broyer par les pompes de brassage). A noter que tous les nudibranches, planaires - vers plats ou plathelminthes - et autres invertťbrťs du mÍme style ne sont pas forcťment parasites contrairement ŗ ce quíon pense souvent. Chelidonura varians, Berghia verrucicornis ou les limaces Tridachia crispata et Elysia sp. (qui "broutent" les algues Bryopsis) en sont de parfaits exemples.



Cheilinodura varians et les planaires.
Photos, Christian SEITZ et Julien THEODULE
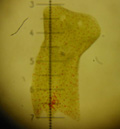
La planaire Convolutriloba retrogemma
Photo, Ph. et L. COLON
D'autres clichťs :
Berghia verrucicornis
Berghia verrucicornis finissant de dťvorer une aiptasie
Tridachia crispata
Du point de vue technique, lorsqu'on utilise une espŤce prťdatrice ou parasite díune autre espŤce qui est considťrťe comme nuisible, la premiŤre espŤce est taxťe d'espŤce "auxiliaire". L'espŤce auxiliaire est utilisťe pour lutter biologiquement contre l'espŤce nuisible.
Le parasitisme
Le parasitisme est un mode de vie oý un individu "parasite" vit aux dťpens d'un individu "hŰte" sans le tuer (sinon on parle plutŰt de prťdation). Ce mode de vie peut Ítre obligatoire dans les sens oý le parasite ne peut pas survivre sans son hŰte [chez le virus : exemple des virus du SIDA (La Lettre de líInstitut Pasteur, 1995)]. Cette relation implique de nombreuses adaptations. La premiŤre est morphologique : le parasite est plus petit que son hŰte car le parasite vit dans son hŰte (endoparasite) ou sur son hŰte (ectoparasite). Il y a ťgalement des adaptations physiologiques : dťveloppement díorganes pour s'agripper ŗ líhŰte, rťduction des organes locomoteurs, fertilitť trŤs importante (reproduction sexuťe et asexuťe) Ö La formation des binŰmes hŰte/parasite implique aussi une spťcificitť parasitaire : líespŤce parasite ne síattaquera quíŗ peu díespŤces hŰtes diffťrentes. Cela conduit ŗ une diminution de la compťtition entre les parasites de diffťrentes espŤces mais diminue aussi les probabilitťs de rencontrer un hŰte. Ces deux points doivent síťquilibrer pour la survie de líespŤce parasite. Le parasite ne doit pas Ítre trop virulent, et ne doit pas tuer son hŰte trop rapidement, sinon il risque de mourir lui-mÍme. Si le cycle de dťveloppement du parasite ne nťcessite quíun seul hŰte, le cycle est dit monoxŤne, síil y a plusieurs hŰtes successifs, il est dit hťtťroxŤne. Parfois le parasite modifie le comportement de son hŰte pour favoriser son cycle. Par exemple, líArtemia salina si connue est parfois líhŰte intermťdiaire díun cestode ŗ cycle hťtťroxŤne. LíhŰte final de ce cestode est une espŤce díoiseaux. Les artťmias parasitťes ont une couleur orange plus visible et par ailleurs nagent plus en surface, du coup elles sont plus vite consommťes par les oiseaux que les artťmias non infestťes. L'aspect le plus impressionnant rťside souvent dans le mimťtisme trŤs poussť entre le parasite et líhŰte, en particulier chez les nudibranches. Par exemple le nudibranche Phestilla melanobranchia, parasite des Tubastrea, a une coloration trŤs variable mais toujours trŤs ressemblante ŗ celle du Tubastrea parasitť.
Phestilla melanobranchia forme noire
Phestilla melanobranchia forme orange
Phestilla melanobranchia forme jaune

Tubastrea morphe jaune
Photo, Ph. et L. COLON
Cet article a pour but díexposer les concepts scientifiques sur les interactions entre espŤces, avec des exemples pour nos bacs rťcifaux. Pour la partie parasitisme, il y aura une suite plus dťtaillťe sur deux grands parasites qui commencent ŗ devenir prťoccupants : les planaires parasites díAcropora et les nudibranches parasites des Montipora/Porites.

Planaire parasite díAcropora Photo, Ph. et L. COLON

Nudibranche Phestilla melanobranchia parasite de Montipora, adulte avec une ponte
Photo, Ph. et L. COLON
L'amensalisme
Líamensalisme est un type de relation observť le plus souvent observť chez les vťgťtaux, dans lequel une espŤce, en se dťveloppant, inhibe le dťveloppement díautres espŤces. Líamensalisme passe par líutilisation de substances chimiques toxiques pour lutter contre la concurrence par inhibition de la croissance. Par exemple, le noyer Juglans regia produit de la juglone dans les feuilles qui, lorsquíelles tombent, inhibent la pousse des plantes au pied de líarbre (sauf le plantain Plantago lanceolata qui est rťsistant). Autre exemple : líantibiose qui est líinhibition de la croissance par des substances antibiotiques. On parle aussi díallelopathie; ce terme dťfinit plus exactement líexcrťtion de substances toxiques.
Líallťlopathie est connue chez les algues Alexandrium catenella. Cíest une dinoflagellťs toxique, bien connue dans la lagune de Thau oý elle pose rťguliŤrement un problŤme sanitaire. Un laboratoire de Montpellier, "EcosystŤmes Lagunaires", travaille actuellement sur ce cas. Sauf que líťtude de l'allťlopathie en milieu aquatique est difficile in vivo: le phytoplancton dťrive dans le milieu naturel et les substances allťlopathiques sont diluťes [Les substances allťlopathiques de microalgues dťjŗ identifiťes sont des peptides cycliques, alcaloÔdes, acides organiques et acides gras (Legrand et al. 2003)]. Les recherches sur líallťlopathie se sont surtout focalisťes sur les cyanobactťries car celles-ci sťcrŤtent beaucoup de substances bioactives - antibiotiques, antifongiques, antivirales - potentiellement utilisables en biotechnologie et en mťdecine.
MÍme si on parle surtout díamensalisme pour les vťgťtaux, les cas díallťlopathie sont courants, et les luttes entre coraux dans nos bacs en sont un bon exemple. Les Palythoa produisent des toxines qui figurent parmi les plus puissantes de toutes les substances chimiques.
Le commensalisme
Une espŤce est dite commensale lorsque elle vit associťe ŗ une autre espŤce en profitant des dťbris des repas de cette derniŤre par exemple, mais sans lui nuire. Cíest le cas des vers tubicoles logťs dans les porites, de líanťmone commensale Calliactis sp. transportťe sur le dos des bernard líermite.

Vers tubicoles commensaux de Porites.
Photo, Laetitia COLON
Le poisson pilote, espŤce commensale des requins, en est un autre exemple. Dans ce cas on parle aussi de phorťsie, le poisson pilote ťtant transportť par le requin. Cíest díailleurs un cas díťpiphoronte (transport díune autre espŤce ŗ líextťrieur). A líinverse, il existe des cas díendophorontes (transport díune autre espŤce ŗ líintťrieur), par exemple le petit poisson Fierasfer sp. transportť par une holothurie, ŗ líintťrieur du tube digestif et profitant directement des restes de repas de líholothurie. On parle aussi díinquilinisme : le poisson vit dans líholothurie sans la gÍner mais sans rien lui apporter. Il existe aussi des crevettes qui vivent au sein des bťnitiers.
Le saprophytisme
Le saprophytisme est une chaÓne alimentaire ŗ dominance dťtritique : la litiŤre de feuilles mortes est díabord fragmentťe par des insectes de líordre des collemboles, puis reprise par des vers de terre qui forment líhumus, lequel est utilisť par des champignons et minťralisť par des bactťries. Les mÍmes genres de processus se retrouvent dans les lits de sable.
Le mutualisme
Le mutualisme, quant ŗ lui, est une association ŗ bťnťfice rťciproque pour les deux partenaires. On parle souvent de symbiose, mais cíest une notion assez "fourre-tout" : relation dťpendante trŤs forte reliant deux espŤces. Mais avec cette simple dťfinition, un parasitisme est une symbioseÖ Un cas de mutualisme par exemple est celui des figuiers et des guÍpes agaonides. Les agaonides sont des hymťnoptŤres qui pondent dans la figue, milieu favorable au dťveloppement de leurs larves. Se faisant, elles deviennent des pollinisateurs pour le figuier. Il y a aussi líexemple du couple formť par les arbres Cecropia et des chauves-souris frugivores. Ces derniŤres consomment les fruits charnus produits par les Cecropia et ainsi participent ŗ leur dissťmination. Un exemple trŤs courant dans nos bacs est líendosymbiose entre les zooxanthelles (dinoflagellťs) et les cnidaires. Le mutualisme est tel que ces algues Symbiodinium sp. vivent dans les cnidaires eux-mÍmes, dans des cellules endodermiques (cellules supťrieures de la "peau" dit de faÁon imagťe) et en sont dťpendantes pour faire leur propre photosynthŤse (Allemand et al., 1998)(
Localisation des zooxanthelles dans les tissus coralliens). Ce mutualisme nous convient bien car il donne aux coraux contenant des zooxanthelles la capacitť de se maintenir en bac (si l'ťclairage est suffisant bien sŻr). La majoritť des coraux sans zooxanthelles nťcessitent des nourrissages ciblťs et sont quasi intenables en aquarium.
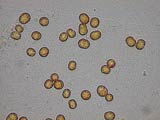
Zooxanthelles
On distingue trois grands types de mutualisme :
Le mutualisme de transport : par exemple des fourmis qui dispersent le pollen ou les graines de la plante. Cíest le cas des fourmis champignonnistes Atta colombica qui coupent des jeunes feuilles de plante et les ramŤnent au nid pour ťlever des champignons dessus. Elles se nourriront ensuite de ces champignons. Lorsquíelles fondent de nouvelles colonies, il arrive que les reines transportent les champignons avec elles, rťalisant ainsi leur dispersion. 90% des plantes dťpendent des vertťbrťs pour la dissťmination (oiseaux, ťlťphants, singesÖ).
Le mutualisme de protection : cíest le plus rťpandu, il peut Ítre facultatif ou obligatoire. Les plantes protŤgent les fourmis en leur fournissant le logement : la domatie, partie intťgrante de la plante et dťdiťe aux fourmis. Ou ŗ líinverse, les fourmis protŤgent la plante de ses herbivores, voire mÍme du feu (crťation díune zone coupe feu au pied des acacias africains en ťliminant systťmatiquement toutes plantes, lianes ŗ proximitť de líarbre hŰte). Certains acariens protŤgent des plantes aussi.
Le mutualisme de nutrition : les Myrmťcodia sont des plantes díAsie tropicale portant des tubercules creux qui abritent les fourmis Iridomyrmex. Les cavitťs lissent servent de loges pour le couvain et les fourmis. Les cavitťs rugueuses servent de zones de dťpŰts pour les matiŤres organiques pour les fourmis et oý la plante plonge ses racines pour se nourrir. Les plantes fournissent aussi souvent ŗ manger aux fourmis en possťdant des nectaires extrafloraux ou des corps nourriciers riches en lipides ou protťines (corps de Belt, corps de MŁller).
Parfois les trois types de mutualisme sont associťs : la plante Myrmecodia est nourrie par les fourmis qui rťsident dans la plante, tout en la dťfendant des herbivores et en dissťminant ses graines, formant ainsi des jardins de fourmis (HŲlldobler et Wilson 1990).
Le mutualisme peut Ítre facultatif, cíest le cas le plus frťquent. Ici, chaque partenaire peut vivre sans líautre et cette association rassemble souvent plus de deux partenaires. Une espŤce particuliŤre peut Ítre associťe ŗ diverses espŤces assez proches pouvant donc rťaliser les mÍmes interactions. Ces espŤces partagent ŗ peu prŤs la mÍme niche ťcologique (pour rťsumer la notion de niche ťcologique : environnement dťpendant de variables du milieu, tempťrature, humiditť, nourritureÖ, dans lequel des espŤces peuvent vivre et se reproduire), cíest une guilde díespŤce, et elles sont interchangeables du point de vue de leur rŰle dans líinteraction.
Le mutualisme peut aussi Ítre obligatoire. Dans ce cas, une espŤce ne peut vivre sans líautre, et ce type de mutualisme aboutit ŗ une coťvolution des deux espŤces. La coťvolution, selon Janzen (1980), est une ťvolution rťciproque de deux espŤces A et B : líespŤce A ťvolue ŗ cause díune pression de sťlection exercťe par líespŤce B et, ŗ cause de líťvolution de A, il y a une pression de sťlection sur líespŤce B et celle-ci ťvolue. On peut aussi dťterminer une coťvolution diffuse dans le cas díinteraction ťvolutive entre deux guildes díespŤces. Cíest díailleurs la coťvolution diffuse qui est la plus rťpandue dans la nature. Les systŤmes bactťrie/hŰte en sont un bon exemple : les bactťries Rhizobium et les lťgumineuses telle la luzerne Medicago sativa. Les bactťries vivent dans les nodules de la plante et sont nourries par la plante. En ťchange, elles permettent ŗ la luzerne de fixer líazote atmosphťrique (ce que les autres plantes ne peuvent pas faire).
Líevolution et la coťvolution sont un vťritable terrain de jeu pour les chercheurs. Si vous vous souvenez d'Alice au Pays des Merveilles, vous devez vous rappeler quíavec la Reine Rouge, il faut toujours courir. On court mais on níavance pas, on court en fait pour rester sur place. Du point de vue de l'ťvolution, cíest la mÍme chose, cíest une course perpťtuelle entre les proies et les prťdateurs, entre les hŰtes et les parasites. Le parasite ťvolue et devient trop virulent, du coup líespŤce hŰte doit ťvoluer si elle ne veut pas disparaÓtre. Et ainsi de suite, le parasite doit ŗ nouveau ťvoluer en rťponse ŗ líťvolution de líespŤce hŰte. Tout le monde ťvolue, tout le monde court, mais au final les interactions níťvoluent pas, cela revient ŗ faire du surplace : cíest ce qui a donnť naissance ŗ la Thťorie de la Reine Rouge. Les biologistes sont de grands enfants finalement Ö
Toutes ces diffťrentes interactions sont plus diversifiťes et plus intenses en milieu tropical que dans les milieux au climat tempťrť, car les conditions abiotiques (les facteurs physicochimiques : composition de líeau, tempťrature, pH, perturbations Ö) ne constituent pas un frein aux organismes ŗ cycle court. Si on reprend l'analogie avec la Reine Rouge : celle-ci "court plus vite" dans les ťcosystŤmes tropicaux.
Evolution des interactions
Toutes ces interactions interspťcifiques ne sont pas fixes. Selon les conditions abiotiques ou biotiques (les autres organismes de la mÍme espŤce ou non, partenaire sexuel, compťtiteur, prťdateur, parasite, symbiote) du milieu, le mutualisme peut basculer vers le parasitisme et vice versa. Un exemple frappant de ce basculement est celui de deux oiseaux des forÍts tropicales díAmťrique Centrale. LíOropendula construit son nid alors que le Molothrus, comme notre coucou europťen, níen construit pas. Ce dernier pond donc ses úufs dans le nid díautres espŤces, et ses petits sont ťlevťs par les Oropendula au dťtriment de leur propre couvťe. Cette interaction est donc un parasitisme. Cependant, dans certaines rťgions, il y a des diptŤres parasites qui font díťnormes ravages sur les couvťes. Mais lorsque le nid abrite un Molothrus, les pertes dues aux diptŤres deviennent nťgligeables car les poussins de Molothrus les mangent. Dans cet environnement particulier, le parasitisme devient une interaction ŗ bťnťfices rťciproques : un mutualisme (Smith, 1968). Il existe díautres exemples de glissement entre mutualisme et parasitisme, conditionnť par les variations du milieu, mais qui sont moins radicaux : les buffles africains sont souvent accompagnťs díoiseaux qui leur enlŤvent les tiques et autres parasites. Ils se nourrissent tout en prťservant líhygiŤne de leurs hŰtes. Cette association peut devenir nťfaste lorsque les oiseaux recherchent leur nourriture dans les plaies et les aggravent.
Dťtermination des interactions
Ce quíil faut ajouter, cíest que le classement en association bťnťfique ou nťfaste ne se fait pas que sur de simples observations de comportement. Pour mesurer les coŻts et bťnťfices en biologie, on se rťfŤre ŗ la "valeur sťlective" ou "fitness" díun individu. Terme cher aux scientifiques, de dťfinition un peu dťlicate, on peut le concevoir comme la mesure du succŤs díun individu en tenant compte de tous les paramŤtres possibles : succŤs reproducteur, taux de croissance, survie, rťsistance aux parasites, maladies, etc Ö Les cas citťs plus haut sont des cas "validťs" de mutualismes. Beaucoup de cas souvent dťcrits comme des symbioses dans le langage courant ne sont pas aussi nets. Le cas du labre nettoyeur et des poissons nettoyťs en est un exemple parfait. Une rťcente ťtude anglaise a portť sur un couple de demoiselles ayant sur son territoire, ‚prement dťfendu, une station de nettoyage de Labroides. La fameuse "fitness" de ces demoiselles a ťtť ťvaluťe et le rťsultat est quíelle níťtait pas significativement diffťrente díun couple de demoiselle qui n'aurait pas de labres nettoyeurs sur son territoire. Donc mÍme síil semble ťvident ŗ premiŤre vue quíavoir une station de nettoyage sur son territoire est un avantage, líťtude a montrť quíelle níapportait pourtant globalement rien de significativement positif en plus. Le cas des associations clowns/anťmone rentre dans le mÍme schťma et entraÓne encore de nombreuses discussions ŗ ce sujet.
Voir par exemple les articles :
"Les anťmones des poissons clowns" de C. Seitz sur Rťcifs.org
"Les relations symbiotiques" de J. Le Jardinier sur Rťcif France
Les interactions interspťcifiques dans nos bacs
Dans nos bacs, de nombreux partenariats - sans aller jusquíŗ dire que ce sont des mutualismes - sont dťcrits et sont sympathiques ŗ observer. Mais contrairement ŗ la tendance gťnťrale des aquariophiles ŗ acheter du "gros", ce sont souvent des espŤces de petites tailles qui sont concernťes et qu'il faut maintenir.

Couple díA. ocellaris dans une Heteractis.
Photo, Laetitia COLON

Les Pterapogon kauderni avec líoursin Diadema setosum
Photo, "Le Bac de Michel"

Les crevettes pistolets et leurs partenaires de terrier piscicoles, ici A. randalli
Photo, Hervť ROUSSEAU
Beaucoup díAmblyelotris sont impliquťs dans de telles associations mais il ne faut pas oublier les
Cryptocentrus sp., les
Stonogobiops nematodes avec Alpheus randalli, et aussi le superbe mais rare
Lotilia graciliosa avec diverses Alpheus.

Les crabes de porcelaine Neopetrolisthes (ici N. maculatus) vivent dans les anťmones
Photo, Christian SEITZ
Il faut aussi noter les crevettes dťparasiteuses Lysmata debelius, L. amboinensis, L. grabhami, et
Leandrites, et les poissons dťparasiteurs, les labres nettoyeurs dont le fameux Labroides dimidiatus, mais aussi les petits gobies Gobiosoma: G. evelynae, G. oceanops et surtout
G. randalli qui semble mieux síacclimater en aquarium du fait díun rťgime alimentaire non totalement dťpendant du dťparasitage.

Crevette dťparasiteuse Leandrites
Photo, Ph. et L. COLON

A. leucosternon et Labroides dimidiatus
Photo, Christian SEITZ
Il existe des crevettes associťes ŗ des anťmones, et le plus souvent elles sont aussi dťparasiteuses :
Thor amboinensis avec diverses actinies, les Periclimenes dont
P. yucatanicus avec Condylactis gigantea, P. pedersoni avec C. gigantea ou Bartholomea annulata,
P. brevicarpalis avec Cryptodendrum adhaesivum.

Crevette Periclimenes pedersoni dans líanťmone Bartholomea annulata.
Photo, Laetitia COLON
Autres associations :
į Les vers ŗ panaches Serpulidae et les petits pagures symbiotiques Paguritta dans les Porites.

Serpulidae et Paguritta dans Porites
Photo, Ph. et L. COLON
į Les crabes boxeurs : Lybia edmonsoni ou
Lybia tessellata. qui portent des anťmones Bunodeopsis comme des gants de boxe.
į Les petits crabes
Trapezia vivants dans les Pocillopora ou les
Tetralia nigrolineata et Domecia vivants dans les Acropora
į On pourrait aussi rappeler líassociation si bien connue des plongeurs entre les bancs de demoiselles dont
Chromis viridis ou díAnthias et les grands Acropora branchus.
Conclusion - Pourquoi tout ceci ?
Une premiŤre rťponse serait peut-Ítre de dire que les biologistes aiment bien se triturer les neurones. Mais Áa serait un peu facile. En fait, tout ceci permet de se rendre compte que rien níest simple et que peu de choses en biologie sont fixes et dťfinitives. Les grands termes symbiose, parasitisme et mutualisme sont connus mais souvent utilisťs sans distinction. En se penchant sur un cas qui parait simple, líanťmone et son clown, on dťcrŤte que cíest une symbiose ! En y regardant de plus prŤs, ce níest pas si simple. Il ne faut pas oublier que nos bacs sont des ťcosystŤmes ŗ part entiŤre, ni de simples tableaux vivants , ni de jolies ou couteuses collections de "cailloux colorťs" ou de poissons "flashies". Ce sont des assemblages hťtťroclites díÍtres vivants, assemblages pas toujours judicieux díailleurs. Ces Ítres vivants interagissent constamment. En aquarium, on retrouve toutes les interactions inter et intraspťcifiques possibles. En savoir un peu plus sur ces interactions permet parfois de mieux comprendre ce qui se passe dans nos bacs.
Avec tout ceci, vous avez eu un aperÁu de quelques notions de biologie qui concernent de prŤs votre aquarium, et jíespŤre que vous regarderez maintenant votre bac díun úil diffťrent !
Sources et rťfťrences :
www.advancedaquarist.com
www.fishbase.org
www.recif-France.com
http://reefkeeping.com/
www.recifs.org/
www.seaslugforum.net
http://perso.wanadoo.fr/aresub/medecinesubaquatique/dangersfaune/biotox.htm
http://www.garf.org/trever/foreignenvironmen.html
www.sbg.ac.at
www.wetwebmedia.com
www.aquatab.net
www.env.go.jp
www.gla.ac.uk
liboupat2.free.fr
La Lettre de líInstitut Pasteur, 1995. 11 : 5-8.
Allemand, D., Furia, P., et Benazet-Tambuttť, S., 1998. Mechanisms of carbon acquisition for endosymbiont photosynthesis in Anthozoa. National Research Council Canada. 76 : 925-941.
HŲlldobler B. et Wilson, O. 1990. The ants. The Belknap Press of Harvard Univ. Press, Cambridge, Mass. 732p.
Janzen, D. H., 1980. When is it coevolution ? Evolution. 34 : 611-612.
Smith, N. G., 1968. The advantage of being parasitized. Nature. 219 : 690-694.
Graf, J. D. et Polls Pelaz, M., 1989. Evolutionary genetics of the Rana esculenta complex.
Evolution and Ecology of Unisexual Vertebrates. 466 : 289-301.
Van Valen, L., 1973. A new evolutionary law. Evol. Theory. 1: 1-30.
Baensch, Hans A., Debelius, Helmut, 1998. Atlas de líaquarium marin. Mergus Verlag. 1215p.
Sprung, Julian et Delbeck, J. Charles, 1999. Líaquarium rťcifal, tome 1 et 2. Ricordea Publishing. 544p et 546p.
Cours de DEUG BBC Avignon Ė licence Bio des organismes Montpellier Ė MaÓtrise Bio des populations Montpellier Ė DEA AMSB Lyon
Pour la plupart des informations sur les problťmatiques mutualisme/parasitisme, les axes de recherche de líťquipe "Coťvolution" du CEFE-CNRS de Montpellier (fourmis figuiers ...) et le laboratoire HydrosystŤmes Fluviaux de Lyon (grenouille verte)
Et tellement de discussions avec des aquariophiles passionnťs Ö !
Article ťcrit par Laetitia Colon, et publiť par Rťcifs.org le 03/08/2005
|
|
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}